In the South American jungle, a jaguar hunts a prey that bites back, a caiman. In a show of absolute superiority, the cat makes his kill on the dangerous dinner’s turf. The jag jumps into the river and snatches the wriggling toothy reptile by its neck.

A caiman or a gator may be fast, but if he misses, he is in deep trouble, as he just shot a musket—a single-shot muzzle loader against a warm-blooded predator’s six-shooter that can be rapidly reloaded. A reptile’s weak aerobic system and reliance on glycolysis puts it at a great disadvantage against warm-blooded predators whose aerobic systems enable them to rapidly recover from their lightning strikes.
A gator needs many hours of rest and sucking wind to clear the acid produced by a single spurt of activity. Scientists believe that dinosaurs—reptiles’ cousins—had similar metabolism, which made them vulnerable to mammal predators whose aerobic systems were far superior and that enabled them to sustain their attacks.
Enter the home of the aerobic system, the mitochondria, our cells’ power plants.

This Quick and the Dead did not start out as a power-centric guide to total fitness with a health twist. The original intention was to write only about improving endurance for high-power applications by training the mitochondria.
As it turns out, the state of one’s mitochondria determines much more than the outcome of a rowing race or a judo match. The mitochondria are, in Dr. Nick Lane’s words, “the masters of life and death.”
The alternative of being “quick” to being “dead” in the African savannah or the old American West only scratches the surface of the inspiration behind the book’s title.
“Who shall give account to Him who is ready to judge the quick and the dead.” ~ 1 Peter 4:5
In the Bible, the word “quick” does not describe being quick on the draw. It means “alive.” The goal of the Q&D regimen is not just to get you to perform at a high level, but to do it in a way that does no harm to your health—and hopefully improves it a great deal.
The Old English word “quicken” meant “to come to life.” The unborn child’s first movement in the womb was called “quickening.” Fittingly, we get our mitochondrial genes from our mothers.
Healthy, strong, and abundant mitochondria make one much more resilient to a variety of stressors: cold, heat, altitude, infection, poison, radiation, etc.
On the other hand, mitochondrial dysfunction is a likely cause of cardiovascular and neurodegenerative diseases, cancer, diabetes, obesity, and aging.
Oftentimes the events leading up to these disasters involve free radical damage. Reactive oxygen species or free radicals are oxygen molecules that carry more electrons than they are supposed to. Like lactic acid, in moderate quantities, ROS perform many useful functions. In excess, they destroy performance, health, and life itself.
Mitochondria are the primary source of free radical generation in our bodies. When there is heavy traffic through the so-called electron transport chains in mitochondria, some of the electrons leak and get captured by oxygen molecules. The latter turn into highly reactive compounds like hydrogen peroxide that go on a rampage damaging the body.
You can eat blueberries until you are blue in the face, yet they are not going to save you from ROS: Antioxidant foods and supplements have turned out to be surprisingly ineffective[i] (they might even increase cancer risk.[ii])

What you need are more, bigger, and better mitochondria.[iii] They not only generate and leak fewer free radicals—they act as “net sinks,” as Dr. Alexander Andreyev put it.[iv]
If you plan on remaining quick—as in “alive”—as long as possible, you had better treat your mitochondria right.
Choices that damage mitochondria include the usual suspects of smoking, drinking, overeating, eating garbage, pollution, stress, and overtraining.
Choices that beef up mitochondria are caloric restriction, intermittent fasting, controlled hypoxia and hypothermia—and certain types of exercise.
Scientists have identified four primary signals that can lead to mitochondrial biogenesis or creation[v]:
- Mechanical strain
- Reactive oxygen species
- Increased calcium concentration
- Low energy status
One at a time.
It appears that classic Soviet anti-glycolytic training—“A+A” on our site—builds mitochondria in fast twitch fibers through the first mechanism.
The second demands “optimal”[vi] or “physiologic”[vii] concentration of free radicals. It seems to apply to intelligent and restrained interval training—as opposed to acid baths like “Tabatas”.
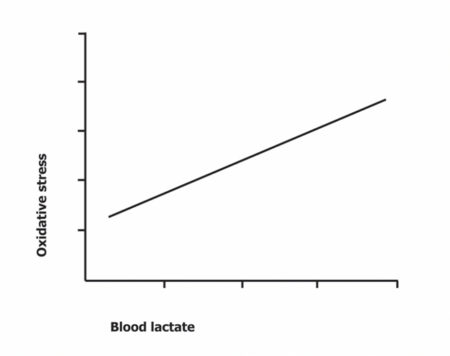
“Metcons” are custom made for maximizing ROS generation. You may have heard of glutathione (GSH), “the mother of all antioxidants” produced by our bodies. Scientists measure the concentrations of its “used up” version to assess oxidative stress. They found a linear relationship between its concentration and lactate concentration after exhaustive exercise[viii].
The third mechanism works only in slow twitch fibers.
The fourth is what “high intensity interval training” attempts to harness.
This mechanism was discovered by preeminent Soviet scientist Prof. Felix Meerson[ix] back when Deep Purple was recording Speed King. Unfortunately, Meerson’s discovery was overlooked and had to be made again almost half a century later.
In the XXI century, Western scientists did a very fine job of rediscovery—going beyond the Soviet professor in tracing the specific metabolic pathways starting with a low energy state and arriving at mitochondrial biogenesis or creation. Alas, other scientists, those who experimented with HIIT, paid token attention to these discoveries and viewed them out of context of the many other biochemical reactions taking place at the same time and some of the events happening in the body at large. The resulting training protocols worked, but often inefficiently and with serious side effects. Like drugs with long disclaimers in small print.
Deep soreness. Low energy. Stress. Hormones out of whack. Free radical damage. Unfavorable changes in the heart.
I became convinced there was a better way.
The Quick and the Dead: Total Training for the Advanced Minimalist

References
[i] Teixeira et al., 2009
[ii] Yun & Finkel, 2014
[iii] Hood, 2001; Boveris & Chance, 1973; Davies et al., 1981; Jenkins et al., 1983
[iv] Andreyev et al., 2004
[v] Coffey & Hawley, 2007
[vi] Lira et al., 2010
[vii] Paulsen et al., 2014
[viii] Sastre et al., 1992
[ix] Meerson, 1971




I am in my mid 30’s and already doing Kettlebell S & S. Should I try Q&D ? How is it different from S & S?
Nahid, very different. I suggest that you accomplish the Simple goal and excel in pushups before considering Q&D.
Been reading Pavel since the last 90s and I’m excited about this new release. I have a fresh highlighter and notebook ready.
I apologize if this is too off topic, but I’m following the Q&D protocal (love it!) and the high rate and magnitude of creatine phosphate depletion, coupled with minimizing glycolysis, makes me curious if it dovetails with another breath-orientated protocol I’ve been following, which was laid out by Wim Hof “The Iceman”.
Basically, Wim has me deeply and rapidly breath 30x, exhale fully, then hold my breath for as long as possible. I do 4 rounds. The connection with the Q&D occurs on the fourth round, where I do as many push-ups as possible while holding my breath after exhaling.
Do you think Wim has stumbled upon a way to rapidly deplete CP while avoiding glycolysis altogether, due to my body being over-oxygenated? Interestingly, most people can do significantly more pushups in the 4th round vs in a normal situation. For example, normally I can only do around 40 push-ups in a row (I know, I’m working on it!). However, after 4 rounds of rapid breathing, breathing all the air out, then doing pushups, my best is 75.
Thank you for any insight into the biochemistry you can offer!
Glenn, hyperventilation makes your system more alkaline which enables you to build up more acid.
That said, there are many many medical warnings against hyperventilation. I discuss this in detail in the Second Wind seminar.
Thank you for taking the time to respond! While the Second Wind seminar seems geared towards athletes, it seems equally powerful from the perspective of general health and extending one’s healthspan. A book covering this topic would be invaluable for the general public 🙂
Nice mention of cold stress!
Cold thermogenesis is arguably one of the best, certainly one of the least used, tools to power up the mitochondria.
Comrade Pavel, thank you for the book excerpts. Please tell me the audible addition is on its way. I’m a totally blind grappler, and your “Simple and Sinister” audiobook helped me get bak into competition shape; after three years off.
Jonathan, the audio book is coming. Power to you!
After reading the book, the conclusion is quite simple: it is impossible to overstate the value of the Quick and the Dead, both as a scientific paper and as a reading experience. One question remains, however.
I am aware that S&S and Q&D can be alternated to great effect. Furthermore, it is clear that (as Pavel’s answer to Bauer suggest) that one should not be mixed with the other. Apples and oranges are to be kept separate, no fruit salad is allowed, Comrades. Is it possible, however, to have one after the other? Fast first, says the credo, and it is as logical as simple. But can one, after completing his daily dose of Q&D and presuming sufficient recovery after it, do S&S? In chapter ‘The Schedule’ Pavel writes that Q&D’s nature makes it easy to fit with other types of training, yet (thus far) I have seen no reference to having S&S after it, only alteration and combination.
Can, or rather, should one eat apples after oranges? Can one go and practice with the Glock at the range after hunting gazelles?
Thank you kindly for your answer in advance.
Leopard Power to You!
Balázs Sz.
Balázs, it is not optimal to do both Q&D and S&S at once; there is too much overlap.
Hi Pavel,
Have been checking Amazon for a kindle version but nothing seems to come up.
Is the kindle version on its way soon?
Rob
https://www.amazon.com/Quick-Dead-Training-Advanced-Minimalist-ebook/dp/B07W2WMZGP/
Thanks for the link, Steve.
Much appreciated.
Rob
Com. Pavel, thank you for writing this book, it is truly a masterpiece! Very logical and to the point.
I am still on my way to achieving simple and have a question regarding the Fast 10’s and rest periods: Would you recommend these days to train S+S in a circuit fashion, like “Victor”? Swings every 3 minutes with 1 GU inbetween around the 1:30 mark?
Would this be different for beginners and intermediate trainees, as power production and thus ATP and CP depletion is less pronounced in the former?
Thank you, Bauer.
No, I do not recommend training S&S in a circuit. Stick to the book and abide by the talk test.
Very much looking forward to the audiobook!
Thank you, gents!
References? They seem more like rough suggestions. First one doesn’t exist; second one does not support the statement made. And so on. How about a proper list with names of the papers and the journals? And maybe have them checked before publication. Sorry if this sounds pedantic. But why have useless citations?
Simon, I do not appreciate your attitude but I will make an exception once.
Here is the link to the first reference: https://www.ncbi.nlm.nih.gov/pubmed/19657294
Take your time reading the second one instead of skimming it:
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4016106/
Going forward, roll up your sleeves and track the citations yourself.
Pavel,
This is so interesting and just what I have been finding out through my mistakes and successes in training, sparked on by reading your exposés in your ‘cost of adaptation’ and other landmark blogs, and of course all your books that I own. Really a lifechanger for me after having trained with the lactic mentality for a couple of decades, and always getting sick, always wrecking my joints.
I’m building mitochondria as I write this through my hunger strike protest at the kindle edition being so elusive…can’t wait to read it!
Thanks once again
Rob
Forgive me if I should know this, but is Q&D the published version of Strong Endurance? Can’t wait for the book to arrive!
@Jack Staples,
Consider the relationship of our three-day kettlebell instructor certification workshops to the book, “Kettlebell, Simple and Sinister.” The cert is both wide and deep, discussing principles and how they should be applied, by both students and potential teachers, over a variety of applications; the book contains everything you need to know in order to understand and to execute its minimalist program. The book and that minimalist program have a beauty and simplicity all their own; the three-day workshop is for those wishing to explore the material in much greater detail and to possibly use to train their own students.
The same holds true for the Strong Endurance workshop and the Q&D book. Read the book, and if you want to know more and possibly to teach your students, the seminar is highly recommended.
NB: Strong Endurance is not an instructor certification, but it is nonetheless highly recommended for those who train others.
Steve Freides
Director of Community Engagement for StrongFirst
Thank you for the thorough response Steve!
Happy to help, Jack.
-S-
Very excited for this new book. Pavel has been a great inspiration for me, keeping me strong, fit and ready for duty for nearly two decades.